はじめに
「発酵」とは、広義には有機物質が微生物によって分解される現象を指しますが、人類に有益な化合物(あるいは物質)が生みだされる点で「腐敗」とは区別されます。さらに広い意味では、「人類に有益な化合物(あるいは物質)が微生物によって生みだされる現象」のすべてが発酵に含まれます。したがって、「発酵学」とは人類に役立つ微生物に関する学問であるということができます。研究室名には今でも「さけのとり」のついた「醗」を使い続けていますが、「醗酵学」という文字の中に、「研究室の歴史」と「微生物への期待」を感じていただければ幸いです。ともすれば、古めかしいと思われてしまう研究室名ですが、我々はこの名前に誇りをもっており、これからも新しい「醗酵学」を切り拓いていきたいと考えています。
『研究室の歴史』を読んでいただければわかるように、これまで醗酵学研究室では、時代を先取りする独創性の高い研究を行ってきました。この「伝統」は今でもしっかりと受け継がれています。醗酵学研究室における「不易流行」を追求していくことが私の目標の1つですが、「不易」すなわち時代の新古を超越して不変であるべきところは、研究に対する取り組み方であると考えています。
- 微生物の多様性の中に秘められた「微生物の可能性」を信じること
- 基礎研究と応用研究を分け隔てなく捉えること
- 低分子、タンパク質、遺伝子、細胞などさまざまな対象をさまざまな手法を駆使して取り扱うこと
- 他人の物真似は決してやらないこと、実験は徹底的に行うこと
- 自分たちの研究にプライドと責任をもつこと、そして研究を楽しむこと
醗酵学研究室では以上のような研究スタイルを大切にしていきたいと考えています。
一方、「流行」すなわちそのときどきに応じて変化していくべきことの中で、重要に思っていることの1つに「共同研究の推進」があります。近年、ライフサイエンスは非常に大きな発展を遂げており、実験技術の進歩には目を見張るものがありますが、今後のさらなる発展のためには「異分野との融合」が不可欠であると思います。これまでの醗酵学研究室では、できるかぎり「自前」で実験を行うことを重要視してきましたが、今後は異分野の研究者(異なった技術・知識をもった研究者)と積極的に連携することによって、新たな研究領域を開拓していく必要があると考えています。
トップページに示した主な研究テーマについて、以下、順に解説したいと思います。研究の概略がわかっていただければ幸いです。
1. 放線菌の形態分化•二次代謝の分子機構とその制御システムに関する研究
生物はさまざまな物質をつくります。このうち、糖、アミノ酸、脂質、核酸など、多くの生物に共通であり、その生物の生命活動に必須な物質を一次代謝産物と呼びます。一方、一次代謝産物からつくり出され、その生物の生命活動に必須でない独特な物質もあり、これを二次代謝産物と呼びます。抗生物質や色素といったものがその代表例です。人類は昔から、これらの二次代謝産物を薬・香料・染料などの生物資源として利用してきました。
土壌細菌である放線菌は抗生物質などさまざまな有用な二次代謝産物をつくり出すことで知られています。我々はその生産のしくみを明らかにし、二次代謝産物の増産や、新たな二次代謝産物の生産に役立てたいと考えています。また、放線菌はバクテリアの一種でありながら、菌糸状に生育し胞子を形成するという複雑な形態分化を行います(図1参照)。このため、放線菌は細胞分化研究におけるモデル微生物として、基礎研究においても重要な研究対象となっています。
醗酵学研究室では、長年、ストレプトマイシン生産菌であるStreptomyces griseusの形態分化・二次代謝を誘導する微生物ホルモン「A-ファクター」に関する研究を行ってきました。2008年には他の研究グループとの共同研究により、S. griseusの全ゲノム配列を決定し、その後、トランスクリプトーム解析、ChIP-seq解析、プロテオーム解析等の網羅的研究を展開しています。このような「網羅的解析」と「個別の遺伝子やタンパク質の精密な機能解析」を両輪として、「新しい概念」を打ち立てられるような研究を目指してきました。遺伝子発現制御機構の巧妙さ、微生物の生存戦略のしたたかさ・柔軟さについては驚かされることが多く、興味は尽きません。
Streptomyces属放線菌は、土壌中のメジャーな放線菌ですが、近年、新たな二次代謝産物の生産菌として、どちらかというとマイナーな部類に属する放線菌である「希少放線菌」が注目されるようになってきました。
我々は2007年から、希少放線菌の1つであるActinoplanes missouriensisの研究に取り組んでいます。A. missouriensisは多数の胞子を内包した胞子嚢を基底菌糸上に形成しますが、湿潤した環境におかれると胞子嚢の外皮が破れ、数十本のべん毛をもつ遊走子が泳ぎだします。遊走子は走化性をもち、極めて速い速度で水中を泳ぎ周り、発芽に適した場所まで移動すると考えられます(図2参照)。
このようにA. missouriensisは「最も複雑な生活環をもつバクテリア」の1つですが、その分子機構についてはこれまで全くわかっていませんでした。我々は他の研究グループと共同でA. missouriensisの全ゲノム配列を決定するとともに、独自にA. missouriensisの遺伝子破壊法や形質転換法を開発し、以下に述べるように、この複雑な形態分化を担う分子機構の解明を進めています。
(1)栄養増殖を行っている菌糸は栄養の枯渇や乾燥状態などいくつかの必要な条件がそろったことを感知すると、増殖から胞子嚢の形成へと舵を切ります。この際、胞子嚢形成に関わる多数の遺伝子の発現が、それぞれの必要な時期に必要な量だけ厳密に制御されて開始されます。このような周辺環境の変化に応じて胞子嚢形成に関わる遺伝子の発現がどのような機構で制御されているか、またそれらの遺伝子がどのような機能を持っているか研究を進めています。(参照 リンク1)
(2)胞子嚢は休眠状態の胞子が多数包まれた「細胞のゆりかご」とも言える構造体ですが、細胞分裂により増殖する単細胞の生物が多くを占めるバクテリアにおいて非常に珍しい多細胞状態の構造体です。このような珍しい構造体は、これまでに知られていない未知の構成因子からなると期待され、胞子嚢全体を包む外皮や、胞子と胞子の隙間を満たすマトリクスなど、胞子嚢だからこそ存在する物質に注目して研究しています。
(3)上述のように胞子嚢は休眠耐久状態にある胞子の生存維持に特化した構造体ですが、周辺環境が乾燥した状態から湿潤した状態に変化すると、外皮が破れて内部の胞子が放出されます。我々はこのダイナミックな変化を、微生物細胞が休眠状態から覚醒する過程を解明するための格好のモデルと捉え、これまでの他のモデル生物の研究では知られていなかった新たな概念を打ち立てるべく、その分子メカニズムの解明を進めています。
(4)胞子嚢から放出された胞子は表層にべん毛と線毛を持っており、湿潤した環境中でこのべん毛を使って非常に高速で泳ぐ遊走子となります。この遊走子は走化性を示し、栄養豊富な環境では運動をやめて発芽し、再び菌糸として増殖を始めることがわかっていますが、これらの過程においても、「遊走子はなぜ高速で泳ぐことができるのか」、「どのようにしてべん毛運動にブレーキをかけるのか」等、未解明の興味深い現象があり、その解明を目指しています。遊走子線毛が物質への付着に関与することも明らかにしました。(参照 リンク2)
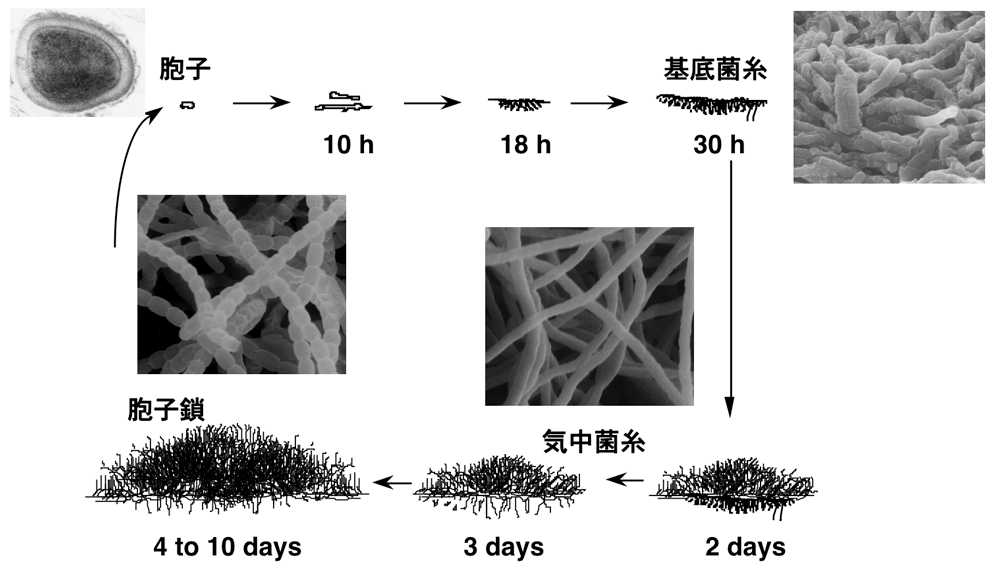
図1. 放線菌Streptomyces griseusの生活環
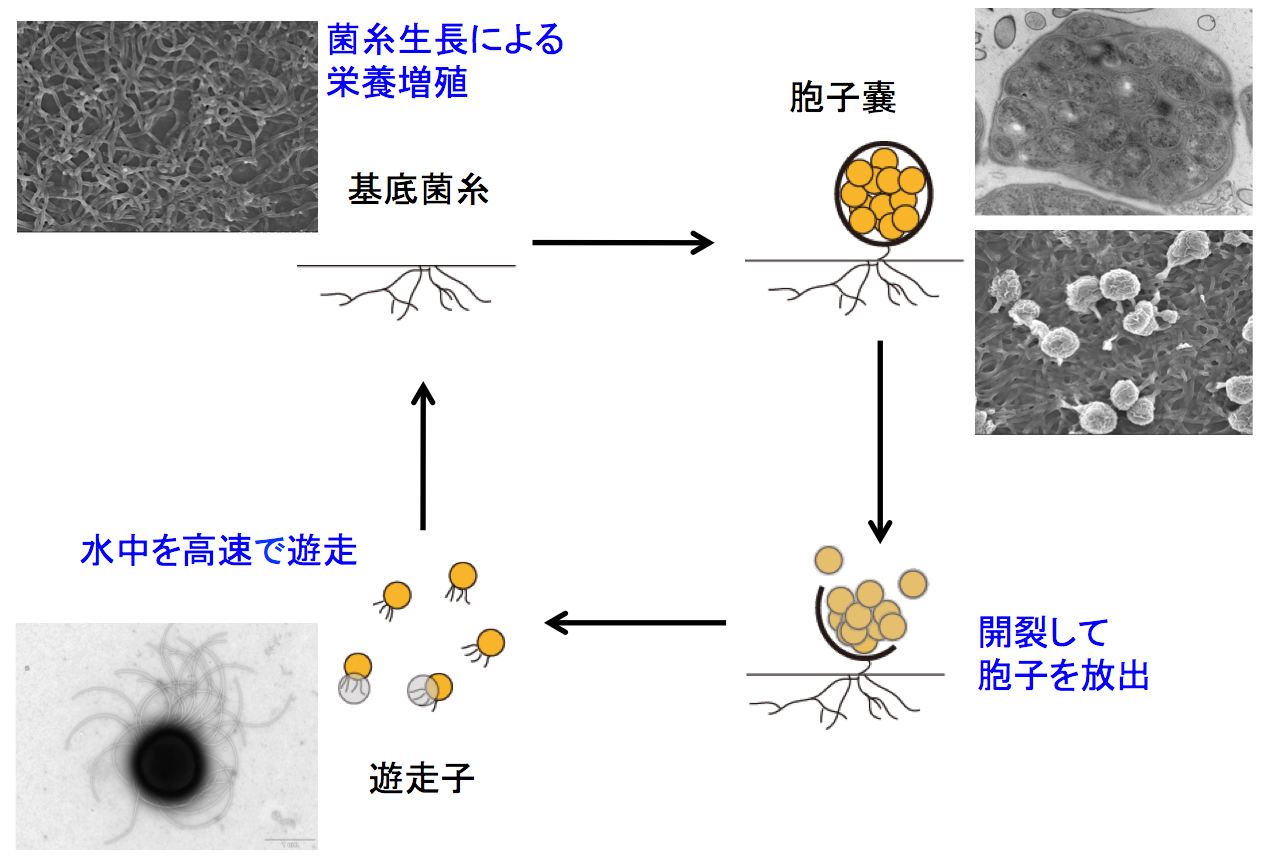 図2. 希少放線菌Actinoplanes missouriensisの生活環
図2. 希少放線菌Actinoplanes missouriensisの生活環
【関連する最新の研究成果】
リンク1 注目を集める希少放線菌の不思議に迫る研究
リンク2 注目を集める希少放線菌の不思議に迫る研究:遊走子線毛の発見と機能解明
2. 微生物の二次代謝産物の生合成に関する研究とその応用
抗生物質をはじめとした多くの「クスリ」が微生物から見出され利用されてきています。これらの「クスリ」は微生物の二次代謝によって作り出される(生合成される)化合物、すなわち二次代謝産物です。二次代謝産物のほとんどは独特な化学構造をもち、多様性に富んでいます。二次代謝産物のこれらの性質が微生物から「クスリ」が多く見出される理由です。これらの独特な化学構造は化合物を作り出すための様々な生合成酵素によって生みだされます。生合成酵素は言うなれば、物質生産のための「専用の分子機械」であると言えます。
遺伝子工学の発展に伴い、種々の生合成酵素を人為的に改変したり新しい組み合わせで利用したりすることによって、新規化合物を生産する微生物を「創り出す」ことが可能になってきています。このような技術は合成生物学とも呼ばれ環境に優しいものづくり実現のための新しい技術として注目を集めています。微生物を「生産工場」として、新しいクスリやポリマー原料などの有用化合物を作るためには、「機械」である生合成酵素の「性能と品揃えの豊富さ」が重要であり、そのためには、新規な生合成酵素の取得とその機能の解明が求められます。
私たちは「生産工場」としての微生物の可能性を追求すべく「専用の分子機械」である生合成酵素を微生物の二次代謝から新たに探し出し、さらにその分子機構を理解する研究を行っています。また、見出した生合成酵素を利用することで、有用物質生産のための新たな「生産工場」を作ることにも挑戦しています。
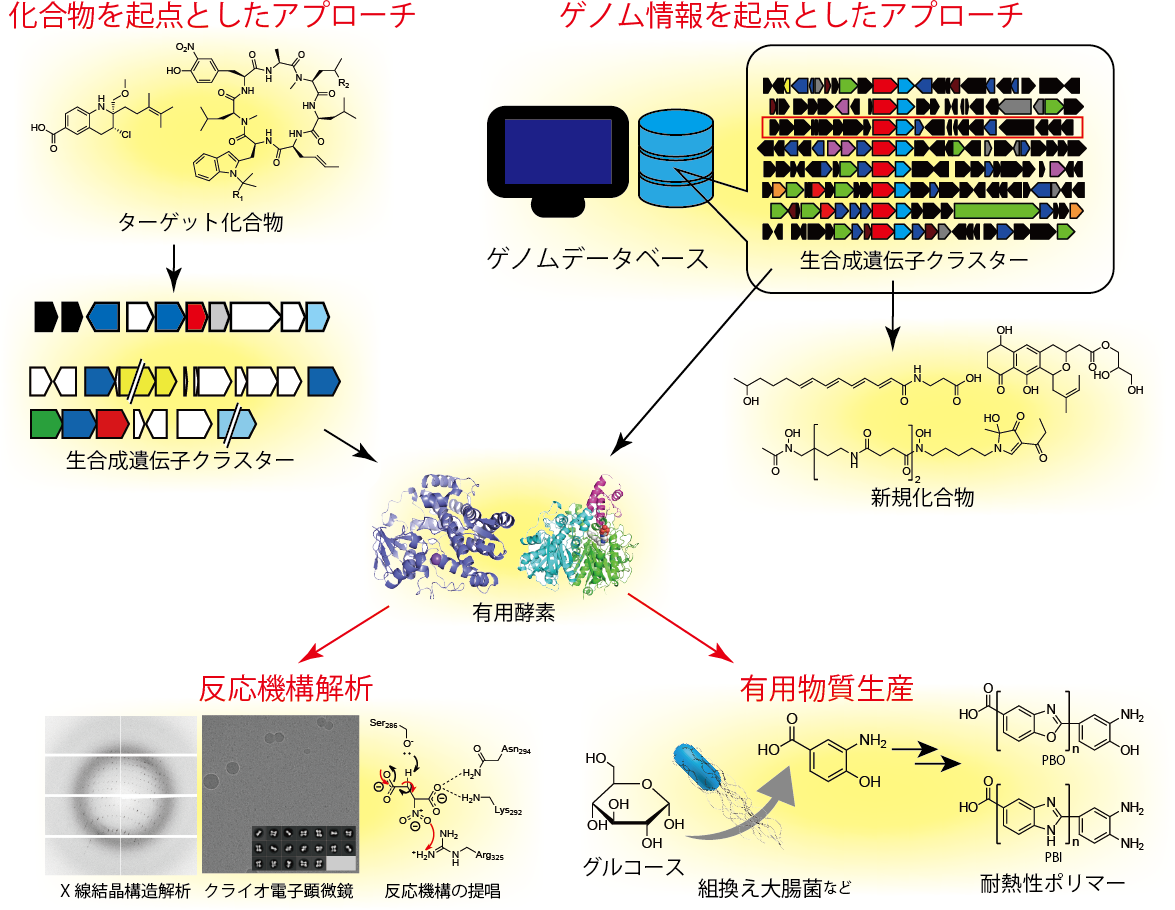
生合成研究全体のイメージ図
2-1 新しい生合成酵素の探索
生合成酵素を新たに探し出すためには2つのアプローチを用いています。1つ目は「化合物を起点としたアプローチ」です。ユニークな構造をもつ化合物に着目し、その構造を生みだすのに必要な新規酵素を明らかにすることを目指しています。このアプローチで研究を行うには、どれだけ「おもしろい」化合物を研究対象にできるかが重要になりますが、微生物代謝産物の単離・構造決定を精力的に進めている研究グループと積極的に共同研究を行っています。最近の重要な成果としては、ジアゾ基を作るための亜硝酸生合成酵素群(参照 リンク3)、ジアゾ基合成酵素(参照 リンク4)、チロシンのニトロ化酵素(参照 リンク5)、ニトレン転移を触媒するシトクロムP450(参照 リンク6)などの発見があげられます。
【関連する最新の研究成果】
リンク3 放線菌に広く分布する二次代謝用亜硝酸生合成経路の発見
リンク4 アラゾペプチンの全整合性経路を解明~微生物がジアゾ基をもつアミノ酸を作る方法が明らかに~
リンク5 ニトレン転移反応を触媒する天然物生合成酵素の発見
リンク6 放線菌が持つチロシンニトロ化酵素の発見
2つ目は「ゲノム情報を起点としたアプローチ」です。これは「ゲノムマイニング」とも呼ばれます。近年のゲノム解読技術の進歩は目覚ましく、多くの生物でゲノム配列が明らかにされてきました。放線菌に限定しても数千種でゲノム配列が解読されています。また、これは不思議なことですが、微生物において特定の二次代謝産物を作るための遺伝子はゲノム上の一箇所にまとまって存在することがほとんどです。このような遺伝子の集まりを「生合成遺伝子クラスター」と呼びます。生合成酵素は独特の化学構造をもった化合物を作り出すための「専用の分子機械」ですが、同じタイプの「機械」はそのアミノ酸配列がある程度似ています。これらの性質から微生物がどの程度、二次代謝産物を作る能力を持っているかを予測することが可能になりました。私たちが主に研究対象として扱っている菌は放線菌ですが、ほとんどの放線菌は30種以上の二次代謝産物を作ると予想されます。それにもかかわらず、1つの放線菌から見つかってくる化合物は通常2から3種程度しかありません。つまり、残りの9割近くは見過ごされてきた代謝能力と言えます。私たちはこの見過ごされてきた「生合成遺伝子クラスター」の機能解析によって、新しい化合物や新しい触媒反応をもつ生合成酵素を明らかにすることを目指しています。この研究により新しい芳香族化合物(参照 リンク7)やポリエン誘導体(参照 リンク8)、シデロフォアなどを見出しています。
【関連する最新の研究成果】
リンク7 天然物の多様性を生み出す生合成マシナリー:フォガシン生合成を担うII型ポリケチド合成酵素に見出されたユニークな戦略
リンク8 新しい酵素サブファミリーを提唱:ポリエンを合成するII型ポリケタイド合成酵素
2-2 生合成酵素の分子機構の解明
生合成酵素は「専用の分子機械」と説明しました。先に述べたとおり、最近の技術の発展により「専用の分子機械」に改造を加えたり、組み合わせたりすることで新しい有用な化合物を微生物に作らせる研究が盛んに行われるようになりました。しかし、「専用の分子機械」に改造を加えるためにはその機械のことをよく知る必要があります。私たちは部位特異的変異導入やX線結晶構造解析(参照 リンク9)、クライオ電子顕微鏡解析などの手法を駆使し、酵素のもつ触媒作用の分子機構 (メカニズム) を理解するための研究も行なっています。
【関連する最新の研究成果】
リンク9 II型ポリケチド合成酵素複合体の全体構造を世界に先駆けて解明
2-3 微生物を利用した有用物質生産に関する研究
有用物質生産の対象として、「新しいクスリ (あるいはその原料) やポリマー原料」を大枠として目指しています。少し古い話になりますが、2006年に我々は新しいベンゼン環合成酵素を発見しましたが、この酵素によって生産される3-アミノ-4-ヒドロキシ安息香酸は強度・耐熱性に優れた機能性ポリマーの原料であり、高分子原料の脱石油化という観点からも注目を集めました。また、最近、北陸先端科学技術大学院大学、神戸大学、筑波大学との共同研究で、3-アミノ-4-ヒドロキシ安息香酸を有機化学的に変換して作ることができる3,4-ジアミノ安息香酸と4-アミノ安息香酸(ABA:アラミド繊維原料) の共重合ポリマーが史上最高の耐熱性を有するプラスチックであることを見出し、紙パルプ糖化液から微生物を用いて生産された3-アミノ-4-ヒドロキシ安息香酸と4-アミノ安息香酸を用いて、この耐熱性ポリマーを作れることを示しました(参照 リンク10)。一方、有用物質生産には酵素機能の改良が不可欠であるため、タンパク質機能改変の技術開発にも挑戦しています。